Population genetics: A revision of the theory of coalescence
La teoría de la coalescencia se ha vuelto trascendental dentro de la genética de poblaciones, dado que proporciona un marco conceptual para el estudio de los procesos históricos que han afectado a las poblaciones y que han dado lugar a la distribución actual de la genealogía de genes. Se basa en un modelo matemático de separación de linajes y deriva génica, que permite analizar la variación genética como un proceso estocástico que describe la forma en que los eventos genéticos poblacionales, van hacia atrás en el tiempo hasta un ancestro común. Además las genealogías de genes han mostrado ser de gran utilidad en campos tan diversos de la biología, como la evolución, la biogeografía y la conservación.
Palabras clave: Genética de poblaciones, coalescencia, secuencias de DNA, genealogías de genes.
The theory of coalescence has become crucial within population genetics as it provides a conceptual framework for the study of historical processes that have affected populations and have led to the current distribution of genealogy of genes. Is based on a mathematical model of lineages separation and genetic drift, for analyze genetic variation as a stochastic process that describes how population genetic events, going back in time until a common ancestor. Besides the genealogies of genes have shown to be useful in diverse areas of biology, such as evolution, biogeography and conservation.
Key words: Populations genetics, coalescence, DNA sequences, gene genealogies.
La genética de poblaciones es una disciplina interesada en el origen, cantidad y distribución de la variabilidad genética presente en las poblaciones naturales, así como el destino de esta variación a través del tiempo y el espacio. Esto constituye la base fundamental del cambio evolutivo, por lo que la evolución se considera como la transformación genética de las poblaciones en el tiempo y el espacio (Templeton, 2006).
Uno de los objetivos principales de la genética de poblaciones es entender cómo evolucionan las poblaciones bajo ciertas condiciones. Para ello es importante tomar en cuenta la frecuencia de un “gen mutante” dentro de la población, la cual se calcula considerando solo una muestra de la misma. Las cuestiones biológicas que se originan de la muestra están relacionadas con la historia de la población que le dio origen y/o con las fuerzas evolutivas responsables de las características observadas (Fu y Li, 1999).
Desde la década de 1980, la acumulación de datos procedentes de secuencias de DNA favoreció el desarrollo de la genética de poblaciones y la demostración de los principios que permiten inferir los eventos que ocurrieron en el pasado y que son responsables de la variación actual observada en las poblaciones (Fu y Li, 1999; Nordborg, 2000).
Para determinar los procesos históricos y demográficos de una población a partir de una muestra, se necesitó de un enfoque nuevo (Fu y Li, 1999). Para ello, los genetistas de poblaciones construyeron modelos matemáticos y propusieron métodos para probar e inferir los parámetros de los modelos a partir de los datos moleculares disponibles (Nielsen y Slatkin, 2013).
De este modo, surgió la teoría de la coalescencia como una alternativa para estudiar la variación genética. Esta teoría permite analizar polimorfismos genéticos (formas diferentes de un gen) poniendo a prueba modelos basados en las fuerzas evolutivas (deriva génica, mutación, recombinación, selección natural, entre otras). Esta teoría se basa en la genética de poblaciones, al tomar en cuenta las fuerzas que producen y mantienen la variación genética, pero a diferencia de los procedimientos tradicionales, se apoya en un marco matemático, estadístico y probabilístico (Avise, 2000). Por lo tanto, la teoría de la coalescencia tendrá dos componentes: el primero, corresponde a los procesos estocásticos que conducen al último ancestro común de la muestra, por medio de diferentes modelos genético-poblacionales; y segundo, las propiedades estadísticas de los métodos de inferencia obtenidas de la muestra, que pueden detallar patrones de polimorfismo (Fu y Li, 1999).
Dentro de la “genética de poblaciones matemática” se reconocen dos figuras principales, Malécot y Kimura. Malécot introdujo la noción de rastrear un par de alelos en el tiempo hacia su ancestro común y una medida de variación genética: identidad por descendencia (Malécot, 1941; 1948), que es la idea central de la coalescencia (Wakeley, 2010). Mientras que Kimura propone la teoría neutral de la evolución molecular (Kimura, 1983).
La teoría de la coalescencia relaciona los patrones de ancestria común entre una muestra de genes y el tamaño y la estructura de la población total (Kingman, 1982a, b; Kuhner, 2008), es por ello que el tiempo de coalescencia para una muestra, será el tiempo de coalescencia de toda la población.
Ya que mientras más individuos se agregan a la muestra, es posible acercarse más al tiempo de coalescencia de la población. Debido a la naturaleza estocástica de la muestra, los tiempos de coalescencia para un locus pueden variar entre distintas muestras. Asimismo, resulta esencial integrar varios loci para obtener valores estadísticos más exactos acerca de la distribución de los tiempos de coalescencia (Nordborg, 2000).
La estrategia básica de la teoría es seleccionar n individuos de una generación en particular y trazar su genealogía, la cual se sigue hacia atrás en el tiempo hasta el punto en que se unen o coalescen y por último se hacen predicciones sobre los patrones de variación genética (Kingman, 1982a).
Desde el trabajo de Kingman (1982a, b), los estudios sobre la teoría de la coalescencia se han desarrollado de manera notable y en la actualidad se reconoce como la piedra angular de numerosos análisis estadísticos basados en datos moleculares provenientes de las muestras poblacionales (Fu y Li, 1999).
La utilidad de la teoría de la coalescencia se debe principalmente a tres características: 1) se trata de una teoría basada en una muestra, dado que para el estudio de una población general se toma en cuenta una muestra de individuos de esa población y se considera que una teoría que describe las propiedades de una muestra es más relevante que la teoría de la genética de poblaciones clásica que describe las propiedades de toda la población; 2) se trata de una aproximación altamente eficiente ya que emplea algoritmos en la simulación de muestras poblacionales sujetas a varios modelos evolutivos permitiendo que diversos aspectos de los mismos se examinen numéricamente; y 3) la teoría puede analizar de manera adecuada la gran cantidad de datos moleculares disponibles, tales como secuencias de DNA, las cuales contienen información sobre las relaciones ancestrales de los individuos muestreados (Fu y Li, 1999). Es decir, los procesos de coalescencia garantizan conexiones filogenéticas entre los genotipos de una especie, en este sentido un evento coalescente ocurre cuando dos linajes de moléculas de DNA se fusionan en una sola molécula en algún tiempo en el pasado (Templeton, 2006).
Además, es posible inferir cambios históricos del tamaño poblacional con base en poblaciones actuales y a su vez combinarlos con filogenias y genealogías para determinar si la población ha sido constante, si ha crecido o bien si ha sufrido algún decremento exponencial a través del tiempo. Se ha visto en algunos estudios que el tiempo de coalescencia no solo se ve afectado por el tamaño efectivo de la población (Ne), sino también por las fluctuaciones en el tamaño poblacional, debido a la selección natural y migración (Avise, 2000; Vázquez-Domínguez et al., 2009).
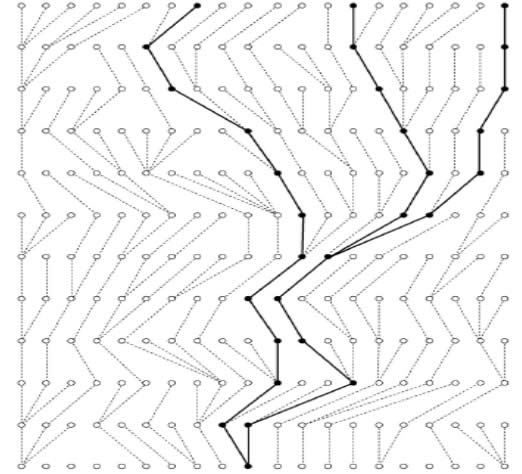
Figura 1. Se muestra una población con 20 copias de genes, donde tres de ellas han sido muestreadas, y los puntos de ancestría común son denominados coalescencias. Tomado de Kuhner, 2008.
Avise, J. C. 2000. Phylogeography: The history and formation of species. Harvard University Press, New York. 447 pp.
Fu, Y.-X. y Li, W.-H. 1999. Coalescing into the 21st Century: An Overview and Prospects of Coalescent Theory. Theoretical Population Biology 56:1-10.
Kimura, M. 1983. The Neutral Theory of Molecular Evolution. Cambridge University Press, Cambridge.
Kingman, J. F. C. 1982a. The coalescent. Stochastic Processes and their Applications 13:235-248.
Kingman, J.F.C. 1982b. On the genealogy of large populations. Journal of Applied Probability 19:27-43.
Kuhner, M. K. 2008. Coalescent genealogy samplers: windows into population history. Trends in Ecology and Evolution 24 (2):86-93.
Malécot, G. 1941. La consanguinité dans une population limitée. Comptes Rendus de l'Académie des Sciences, Paris 222:841-843.
Malécot, G. 1948. Les Mathématiques de l'Hérédité. Masson, Paris. Extended translation: The Mathematics of Heredity. San Francisco: W. H. Freeman 1969.
Nielsen, R. y M. Slatkin. 2013. Inferring Population History and Demography. P. 77-105. En: An Introduction to Population Genetics: Theory and applications. Nielsen, R. y M. Slatkin (eds.). Sinnauer Associates, Sunderland, Massachusetts.
Nordborg, M. 2000. Coalescent Theory. Department of Biological Sciences, University of Southern California. 1-37 pp.
Templeton, A. R. 2006. Populations Genetics and Microevolutionary Theory. Department of Biology. Washington University St. Louis, Missouri. Wiley. 705 pp.
Wakeley, J. 2010. Natural Selection and Coalescent Theory. Harvard University. 1-33 pp.
[a]Egresada de la Licenciatura en Biología y Maestría en Ciencias en Biodiversidad y Conservación, de la Universidad Autónoma del Estado de Hidalgo.